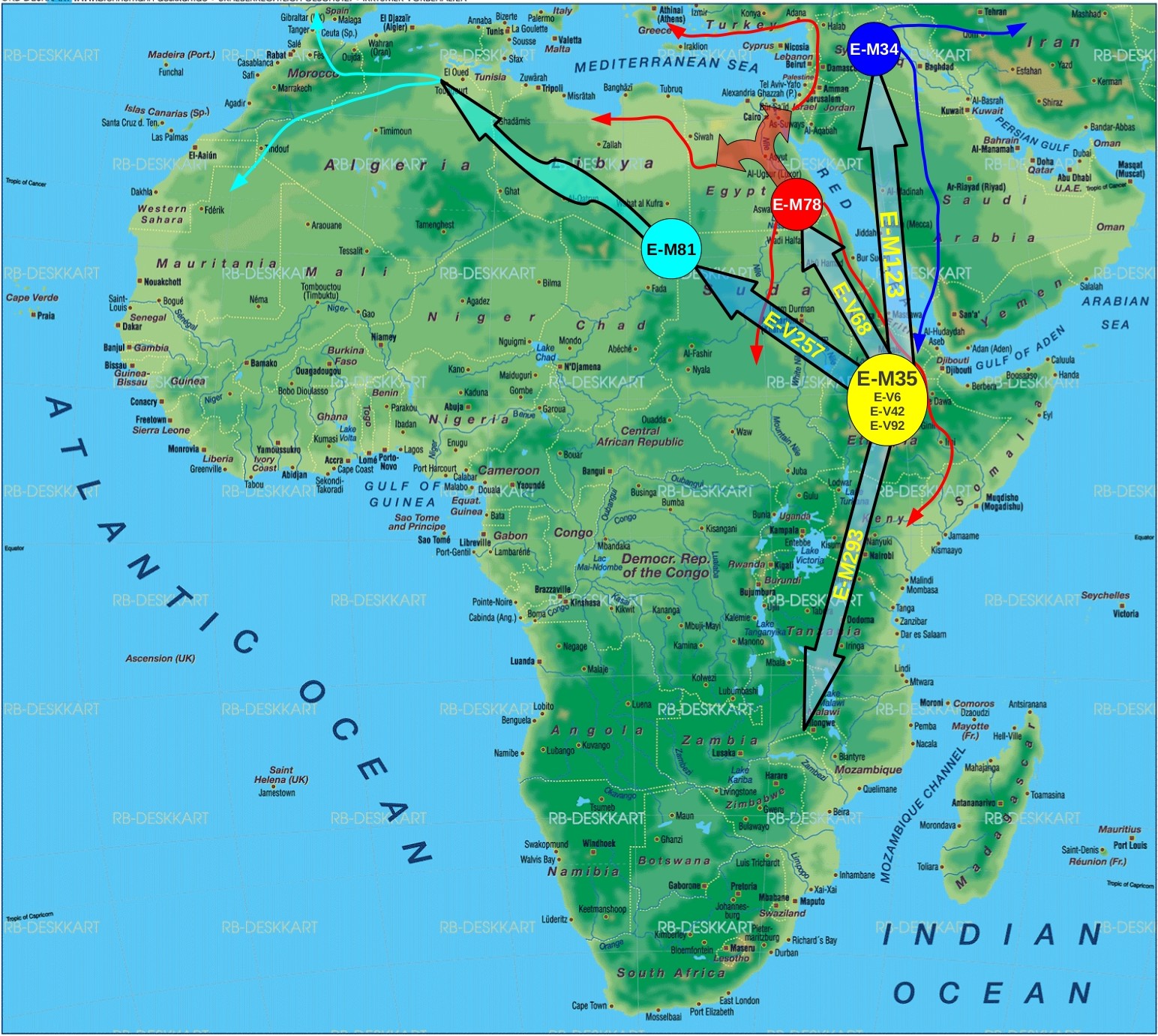
Basically, at first glance, the major novelty with respect to E-M215 is that all E-Z830 (x M123) lineages are united under a new mutation dubbed V1515, and that the former solo lineages of E-M35, i.e. E-V92 and E-V6, now have a home and are included within this unification. In addition, the above named unifying mutation, V1515, apparently has a bifurcated structure itself, with one younger branch having the sole representation in the Southern parts of Ethiopia and further South, and the more diverse (hence ancient) branch being represented in the Northern parts of Ethiopia and further North.
New basal haplogroup E mutations were also apparently found.
The paper is Open access , and I will analyze it further in the coming days , but I just wanted to plot the Eastern African E-M215 variant frequencies for now.

The new fossil calibrated mutation rate has been added to the TMRCA Calulator, unfortunately 95% CI values have not been given (or at least I could not find where they have been given), in any event, central TMRCA estimates for this new mutation rate are a bit slower than mutation rates derived from the other ancient DNA calibrated sources, specifically, ~ 4% and 12% slower than Karmin (2015) and Fu (2014) respectively.
UPDATE (6/27/15) - Comparison with YFull TMRCAs
I have created a table for the TMRCA of the major nodes in E-M215, in order to compare with YFull’s estimates so that we can ‘fill in the gaps’ for the Nodes that have not been given estimates in Trombetta (2015). YFull uses a mutation rate that is almost exactly identical to Fu (2014)’s Ust-Ishim calibrated rates, so naturally some of the TMRCA’s would be closer to today than the Trombetta estimates, as pointed out above.
TMRCA (KYA)
| ||
Trombetta
|
YFull
| |
E-M215
|
39
|
35
|
E-M35
|
25
|
24
|
E-V68
|
20
|
20
|
E-M78
|
15
|
13
|
E-Z827
|
?
|
24
|
E-V257
|
?
|
14
|
E-Z830
|
20
|
19
|
E-M34
|
?
|
15
|
E-V1515
|
12
|
?
|